การแทงผ่าน (Penetration)
การผ่านเข้าสู่พืชดังโครงสร้างและส่วนประกอบของเซลปกติ มีกลไกเกี่ยวข้องอยู่ 2 แบบ
การแทงผ่านเข้าสู่พืชด้วยความดันกล (Mechanical pressure)
การแทงผ่านเข้าสู่พืชนี้เกิดขึ้นจากการเจริญของปรสิต หรือเชื้อเอง พบเกิดจากเชื้อราส่วนมาก ไส้เดือนฝอยและพืชชั้นสูงที่เป็นปรสิต ไม่พบเกิดจากวิสา และบักเตรี ความดันกลที่ใช้บนผิวพืชในการแทงผ่านระดับแตกต่างกันไปขึ้นอยู่กับเชื้อผิวพืช และระดับการอ่อนตัวของผิวพืชจากเอนไซม์ที่เชื้อสร้างขณะที่สัมผัสพืชอยู่ ทำให้เข้าสู่พืชได้โดยทางตรง (direct penetration) การแทงผ่านของเชื้อราและพืชชั้นสูงที่เป็นปรสิต เกิดขึ้นโดยเส้นใยเชื้อหรือรากยึด (rootlet) ยึดติดพืชด้วยแรงระหว่างโมเลกุลที่เกิดขึ้นระหว่างเชื้อหรือรากยึดกับผิวพืชที่สัมผัสกัน เส้นใยหรือรากยึดนั้นจะแผ่เป็นตุ่มเรียกว่า appressorium แล้วเพิ่มเนื้อที่ยึดแน่นมากขึ้น Appressorium จุดสัมผัสยึดพืชมากที่สุดเรียกว่า penetration peg เจริญออกเป็น penetration tube แทงผ่าน cuticle และ/หรือผนังเซล ผนังเซลที่อ่อนการแทงผ่านจะง่าย หากผนังเซลแข็งการแทงผ่านเป็นไปได้ยาก แรงดันที่เกิดที่ penetration peg อาจเป็นความดันออสโมซีส osmotic pressure) ของเชื้อที่เกิดขึ้นนอกเหนือไปจากแรงยึดระหว่างโมเลกุล Penetration tube ซึ่งปกติ มีขนาดเล็กคล้ายเส้นด้ายเมื่อผ่าน cuticle แล้วจะเจริญมีขนาดเพิ่มขึ้นเป็น vesicle คล้ายรูปกรวย การที่ penatration tube เพิ่มขนาดขึ้นนี้อาจเป็นชั้นของ pectin และ cellulose ของผนังเซลมีความต้านทานต่อ การแทงผ่านน้อยกว่าที่ cuticle แล้วเจริญแตกกิ่งเป็นเส้นใยไปตามเนื้อเยื่อต่อไป
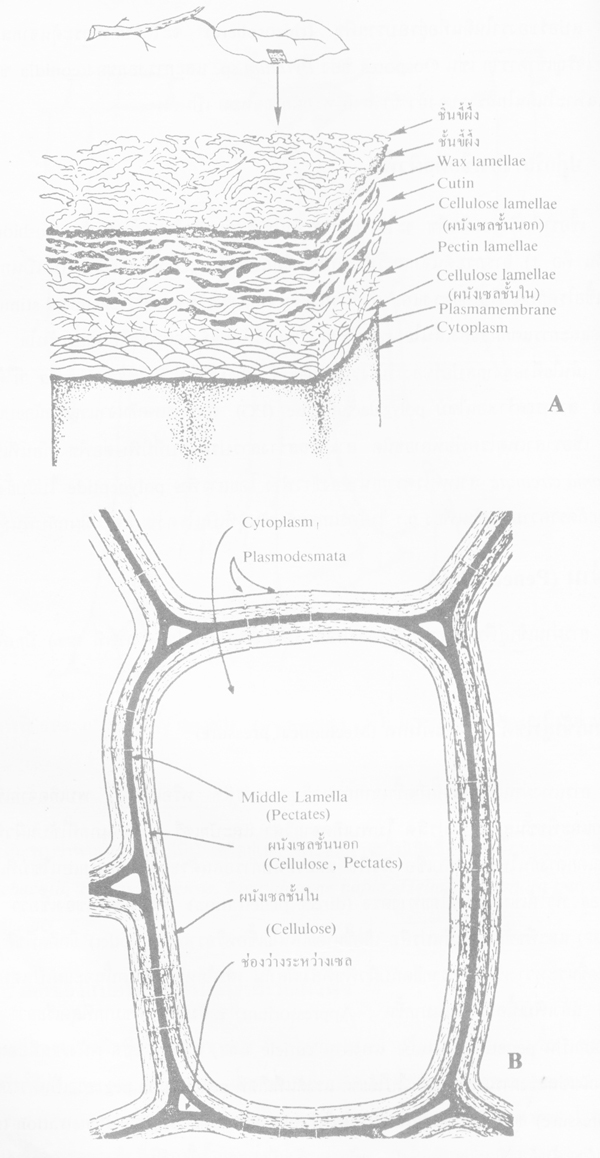
ภาพโครงสร้างส่วนประกอบของเซลพืช A ผนังเซลของใบ และ B เซลทั่วไปภายในพืช(ที่มา: Goodman et al. 1967 และ Agrios, 1978 ตามลำดับ)
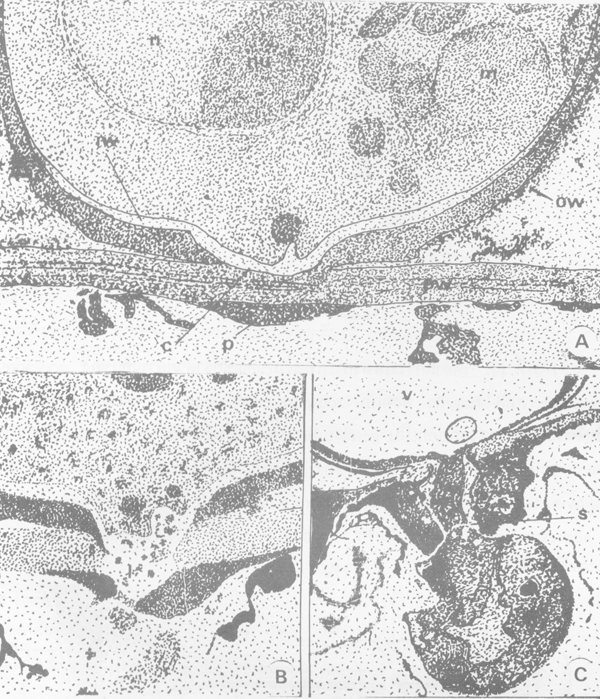
ภาพการแทงผ่านเข้าสู่ใบข้าวโพดของเชื้อ Colletotriohum graminicola A) การเกิด appressorium ในระยะแรก pw = ผนังเซล epidermis C = cuticle,-m = mitochondria,n = mucleus,iw = ผนังด้านใน appressorium ที่สัมผัส cuticle,ow = ผนังภายนอก nu = nucleolus P = papilla ที่เกิดบนผิวด้านในของผนังเซล B) การเกิด penetration peg ตรงกลางจุดสัมผัสของ appressorium และการแทงผ่าน 1 = โครงสร้างคล้าย lomasome ใน infection peg ซึ่งแทงผ่านผนังเซล และ C) การตั้งรกรากของการติดเชื้อ appressorium มี vacuole ขนาดใหญ่อยู่ภายใน(v) และแยกตัวออกจาก infection hypha ด้วยผนังกั้น (s) (ที่มา : Wheeler, 1975)

ภาพไดอาแกรมแสดงการแทงผ่านของเชื้อราด้วยการงอกของ zoospore เข้าสู่พืช โดย zoospore สลัด flagella ออกเป็น zoospore encystment แลว vacuole ภายใน cyst จะขยายใหญ่ขึ้น ในระหว่างการงอกออกเป็น germ tube เพื่อเข้าสู่พืชต่อไป พบเป็นแบบต่างๆ A) Rozella allomycis, B) Olpidium brassicae, C) Chytrium sp., D) Phytophthora parasitica, E) Pythium aphanidermatum และ F) Plasmodiophorales v = vacuole, cw = cyst wall, g = germ tube, Ap = appressorium, Hy = เส้นใย Pt = penetration tube และ Ha = haustorium (ที่มา : Aist,1976)
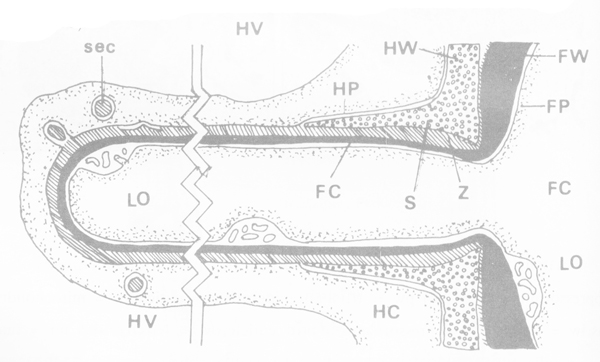
ภาพไดอาแกรมแสดงบริเวณ haustorium ของ รา Peronospora ในพืชอาศัย FW = ผนังเซลของ รา FC = cytoplasm ของ รา FP = plasmamembrane ของ รา Hv = vacuole ของพืชอาศัย HP = plasmamembrane ของพืชอาศัย Z = บริเวณที่แยกระหว่าง plasmamembrane ของเชื้อและพืชอาศัย S = ผนังเซลพืชอาศัยถูกดันตามและคลุมเชื้อ LO = lamasome และ sec = vesicular ของพืชอาศัย (invagination ของเยื่อหุ้มเซลพืชอาศัยเป็นการขับถ่ายอย่างหนึ่ง) (ที่มา : Peyton and Bowen, 1963)
การแทงผ่านผิวพืชของไส้เดือนฝอย โดยไส้เดือนฝอยยึดเกาะพืชด้วยริมฝีปาก แล้วดันตัวเข้าไปในแนวตั้งกับผนังเซล โดยยื่นหลอดดูดอาหารไปข้างหน้าด้วยส่วนหัวที่ยึดติดกับผนังเซลและส่วนท้ายถัดมาจะ หมุนตัวเข้าไปจนเข้าสู่พืชได้ทั้งตัว หลังจากหลอดดูดอาหารแทงผ่านผนังเซลพืช แล้วไส้เดือนฝอยจะขับน้ำลายออกมาละลายผนังเซลช่วยให้การแทงง่ายขึ้น
การแทงผ่านของเชื้อราหรือไส้เดือนฝอยเข้าไปในเซลโดยทั่วไปมีการทำงานของเอนไซม์ร่วมด้วย เช่นย่อยผนังเซล ทำให้อ่อนตัว การแทงผ่านง่ายขึ้น
การแทงผ่านและการทำลายพืชที่เกิดทางเคมี (Chemical attacks)
การเกิดโรคติดเชื้อของพืชเกือบทั้งหมดเป็นผลของปฏิกริยาทางชีวะเคมี ระหว่างสารที่เชื้อปล่อยออกมาในพืชหรือจากที่พืชสร้างขึ้น ส่วนใหญ่เชื้อเป็นผู้สร้างขึ้นและเกี่ยวข้องกับการเกิดโรคทั้งทางตรงและ ทางอ้อม รวมทั้งเอนไซม์ สารพิษ สารควบคุมการเจริญเติบโต (growth regulators) polysaccharides และปฏิชีวนะสาร ซึ่งสารต่างๆ เหล่านี้จะมีความสำคัญต่อความสามารถของเชื้อในการทำให้เกิดโรคมาก แตกต่างกันไป เช่น โรคเน่าเละ เป็นโรคที่เกิดจากปฏิกริยาของเอนไซม์ โรคปุ่มปมเกิดจากปฏิกริยาของสารควบคุมการเจริญเติบโต โรคใบแห้งของข้าวโอ๊ต เกิดจากปฏิกริยาของสารพิษที่เชื้อ Helminthosporium กว้างขึ้น เป็นต้น สำหรับ polysaccharide และปฏิชีวนะสารนั้น มีเชื้อโรคเพียงไม่กี่ชนิดที่สร้างสารนี้ได้ และมักเกิดพร้อมโรคที่เกิดจากเอนไซม์ สารควบคุมการเจริญ และสารพิษ ดังนั้นปฏิชีวนะสาร จึงเป็นเพียงปัจจัยหนึ่งที่ทำให้เกิดโรคได้
เชื้อสาเหตุโรค ยกเว้นวิสาไวรอยด์ และมายโคพลาสมา สามารถสร้างเอนไซม์ สารควบคุมการเจริญเติบโต polysaccharide และสารพิษ เชื้อรา และบักเตรีเท่านั้นที่สร้างปฏิชีวนะสารได้ แต่วิสาอาจทำให้เซลพืชอาศัยนั้นสร้างปฏิชีวนะสารขึ้นในพืช ซึ่งอาจเป็นสารใหม่หรือเป็นสารเช่นเดียวกับในพืชปกติ หรืออาจเป็นสารเช่นเดียวกับที่เชื้อสาเหตุโรคแบบอื่นสร้างได้
เชื้อโรคจะสร้างสารดังกล่าว เมื่อเชื้อเจริญอยู่บนพืชอาศัยหรือในอาหารเลี้ยงเชื้อหรือสารที่เชื้อใช้เป็นอาหารได้ (substrate) สารที่เชื้อสร้างขึ้นอาจไม่เกี่ยวข้องกับความสามารถในการเกิดโรค เพราะว่าใน ปกติก็สามารถสร้างสารนี้ได้ ปฏิกริยาของสารต่างๆ มีดังนี้
1) เอนไซม์ย่อยสารที่เป็นส่วนประกอบของโครงสร้างเซล สารอาหารต่างๆ ในเซล และแทรกระบบการทำงานของพืชตามปกติ
2) สารพิษมีปฏิกริยาโดยตรงต่อ protoplasm และรบกวนความสามารถในการยอมให้ของเหลวผ่านเข้าออกของเซล
3) สารที่ควบคุมการเจริญเติบโตจะมีผลทางด้านโฮโมนในการเพิ่มหรือลดความสามารถของการแบ่งและเติบโตของเซล
4) Polysaccharide ไปรบกวนต่อการเคลื่อนย้ายน้ำในพืช โดยอุดขวางในกลุ่มท่อลำเลียง และอาจมีพิษต่อพืชด้วย และ
5) ปฏิชีวนะสารยังมีการศึกษากันน้อยมาก เข้าใจว่ามีปฏิกริยาเกิดขึ้นคล้ายกับที่เกิดจากสารพิษต่างๆ
เอนไซม์ (Enzymes)
เอนไซม์เป็นโมเลกุลของโปรตีน มีขนาดเส้นผ่าศูนย์กลางประมาณ 2-100 มิลลิไมครอน ทำหน้าที่เป็น catalyst ของปฏิกริยาต่างๆ ในการสร้างโมเลกุลจากโมเลกุลที่เล็กกว่าหรือจากอะตอม และเปลี่ยนแปลงโมเลกุลต่างๆ โดยการเคลื่อนย้ายอะตอมไปสู่ตำแหน่งอื่นๆ ในโมเลกุล หรือแตกตัวให้ไปเป็นส่วนประกอบโมเลกุลที่เล็กกว่า รวมทั้งการทวีจำนวนสารที่ถ่ายทอดพันธุกรรม จากปฏิกริยาดังกล่าว จะได้สารที่เป็นโครงสร้างและพลังงานเพื่อการเกิดสารใหม่ และสังเคราะห์สารโปรตีน เอนไซม์บางชนิดจะมีสารที่ไม่ใช่เอนไซม์ร่วมอยู่ด้วยเป็น coenzymes ในปฏิกริยาเคมีที่เกิดในเซลทั่วไปจะมีเอนไซม์ต่างๆ ที่เป็น catalyst ซึ่งคาดว่ามีเอนไซม์นับหมื่นชนิดที่ประกอบด้วยหลายร้อยล้านโมเลกุล การเกิดเอนไซม์แต่ละชนิดจะถูกควบคุมโดย
หน่วยถ่ายพันธุ์ (gene) หรือสารถ่ายทอดพันธุกรรม ซึ่งขั้นสุดท้ายเป็นกิจกรรมของ mRNA กับ ribosome ด้วยความช่วยเหลือของโมเลกุลของเอนไซม์บางชนิดที่เหมาะสม ส่วนประสิทธิภาพต่างๆ ของเอนไซม์ในการเป็น catalyst ยังไม่ทราบ
เอนไซม์แม้ว่าจะเกิดใน cytoplasm, nucleus, mitochondria และในเยื่อหุ้มของ cytoplasm แต่เอนไซม์อาจสะสมอยู่ในสารที่เป็นส่วนประกอบพื้นฐานของ cytoplasm หรือผนังเซล หรือปล่อยออก
ภายนอก เชื้อโรคที่ต้องการอาหารจากภายนอก อาจปล่อยเอนไซม์ออกมาจำนวนหนึ่ง เพื่อย่อยสารที่ไม่ละลายให้เป็นสารที่เชื้อนำไปใช้ได้ การปล่อยเอนไซม์และสารอื่นๆ ของเชื้อจะมีอิทธิพลต่อกิจกรรมของเอนไซม์ต่างๆ ของเซลพืช
เอนไซม์จำแนกออกได้โดยอาศัยหน้าที่หรือคุณสมบัติเฉพาะออกเป็น 6 แบบ
1. Oxidoreductases เป็นเอนไซม์ที่เกียวข้องกับ oxidation ใน metabolism ของพืช โดยการย้ายอีเล็คตรอน เกิดสารที่เป็น oxidized และสาร reduced
2. Hydrolases เป็นเอนไซม์ที่แตกตัว ester, glycosidic ฯลฯ โดยไปเชื่อมต่อกับโมเลกุลของนํ้าที่ต้องใช้ในปฏิกริยา
3. Lyasis เป็นเอนไซม์แตกตัวที่เชื่อมต่อของ C-C หรือ C-O หรือ C-N
4. Transferases เป็นเอนไซม์ที่ทำให้เกิดการเคลื่อนย้ายกลุ่ม จากสารหนึ่งไปยังสารอื่นๆ
5. Isomerases เป็นเอนไซม์ที่ทำให้เกิดการจัดอะตอมหรือกลุ่มในโมเลกุลใหม่
6. Lygases เป็นเอนไซม์ที่ทำให้เกิดการเชื่อมของสองโมเลกุล โดยใช้พลังงานที่ได้จากการแตกตัวของ energy-rich phosphate bond
การย่อยสารส่วนประกอบผนังเซลของเอนไซม์ (Enzymatic degradation of cell wall substances)
ผนังเซลประกอบด้วย cuticle ซึ่งมี cutin และขี้ผึ้ง (wax) เป็นชั้นคลุมอยู่ และผนัง epidermal cell ซึ่งมี cellulose, pectin และอาจพบสารโปรตีน lignin ปะปนอยู่ด้วย เมื่อเชื้อเข้าสู่พืชแล้วจะเจริญทำลายเนื้อเยื่อ parenchyma ซึ่งผนังเซลประกอบด้วย cellulose, pectin, hemicellulose, middle lamella การย่อยสารต่างๆ เกิดจากเอนไซม์ มีดังนี้
ขี้ผึ้ง ขี้ผึ้งเป็นลิ่มซับซ้อน เรียงกันเป็เนชั้นคลุม cuticle มีช่องที่รอยต่อ ประกอบด้วย primary alcohols เอสเทอร์ของแอลกอฮอล์ และ secondary alcohols, ketones, paraffin hydrocarbons และกรดต่างๆ ขี้ผึ้งแตกต่างไปจากไขมันที่ giycerol ถูกแทนที่ด้วย monohydric alcohols โมเลกุลของขี้ผึ้งส่วนมากประกอบด้วยคาร์บอนด์ 15-37 อะตอม
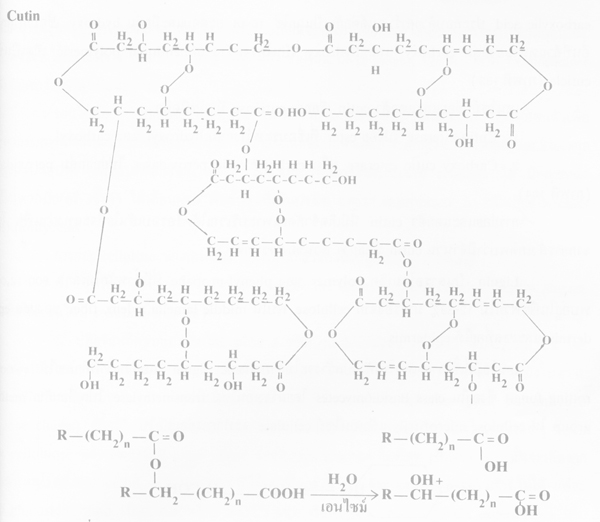
ภาพโครงสร้างของ cutin และการย่อยด้วยเอนไซม์ cutin esterase โดยการ hydrolysis ของ estr bond (ที่มา:Agrios, 1978)
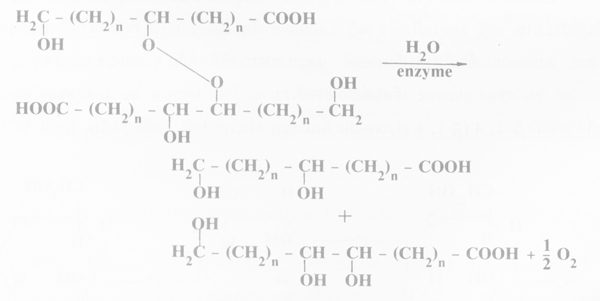
ภาพการย่อยด้วยเอนไซม์ carboxycutin peroxidase โดยแตกตัว peroxide group hydroxy fatty acid 2 chains (ที่มา : Goodman et al. 1967)
เอนไซม์ที่เชื้อสร้างขึ้นและย่อยขี้ผึ้ง (wax degrading enzyme) ยังไม่มีรายงาน แต่ก็เชื่อว่าการย่อยขี้ผึ้งอาจเกิดได้จากจุลินทรีย์ในดินหลังจากพืชตายแล้ว
Cutin เป็นส่วนประกอบหลักของ cuticle เป็น polyester ของ hydroxylated monocarboxylic acid ประกอบด้วยคาร์บอนด์ต่อกันเป็นลูกโซ่ 16-18 อะตอมและมีกลุ่ม hydroxy เชื่อมกับลูกโซ่อื่นที่ตำแหน่งของกลุ่ม carboxy และ hydroxy เป็นโครงสร้าง dimentional polymeric เกิดเป็นชั้น cuticle
เอนไซม์ที่ย่อยและแตกตัว cutin เรียกว่า cutinase มี 2 ชนิดคือ
1. Cutin esterase ไปย่อย ester ที่เชื่อมระหว่างกลุ่ม hydroxyl และ carboxyl
2. Carboxy cutin esterase หรือ carboxy cutin peroxidases ไปย่อยกลุ่ม peroxidase
การย่อยและแตกตัว cutin นี้มีเกิดจำกัดเฉพาะบริเวณใต้หรือรอบเส้นใยที่จะแทงผ่านพืช โดยจากการสังเกตพบว่ามีจำนวน cutin น้อยกว่า ส่วนของพืชปกติ
Lignin เป็นสารอโรมาติค polymer ของ phenyl-propane ที่มีน้ำหนักโมเลกุล 800-12,000 พบอยู่ในพืชพวกไม้ 12-38% มากรองจาก cellulose พบใน middle lamella,xylem, fiber ของเซล epidermis และเซลที่อยู่ใต้ epidermis
เอนไซม์ที่เชื้อราสาเหตุโรคทั่วไปสร้างจะไม่สามารถย่อย lignin นอกจากราที่ทำลายไม้ (wood rotting fungi) ซึ่งอยู่ใน class Basidiomycetes โดยสร้างเอนไซม์ transmethylase ไปเคลื่อนย้าย methyl group ให้ cellulose microfifrils แล้วเอนไซม์ cellulase จะทำการย่อยต่อไป
Cellulose and cellulolytic enzymes
Cellulose เป็นโครงสร้างหลักของผนังเซล มีอยู่ในเนื้อเยื่อของหญ้าประมาณ 12% พืชพวกไม้ที่โตเต็มที่ประมาณ 50% และในฝ้ายถึง 90% Cellulose เป็น microfibril ที่เรียงสานกันประมาณ 280 ถึง 800 สาย ตามแนวยาวด้วยไฮโดรเยนบอนด์ และสภาพทางฟิสิกส์อื่นๆ แต่ละสายจะมีความยาวประม 2000-10,000 หน่วยของ glucose เชื่อมต่อกันยาวประมาณ 1-3 ไมครอน โดยโมเลกุลของ glucose เชื่อมต่อกันที่ตำแหน่ง ß-1, 4 (ß -1, 4 glycosidic linkage) Microfibril บิดยึดกันเป็น fibril ใน fibril ของ
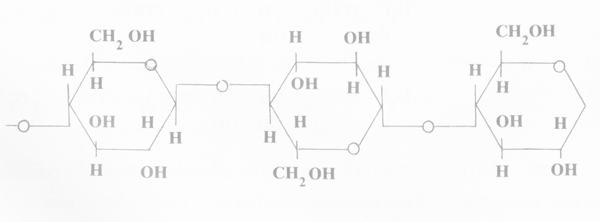
ส่วนที่มี microfibril ยึดสานกันมากก็จะทำให้แน่นและเหนียว บางแห่งที่ยึดสานกันน้อยก็จะหลวมๆ ส่วนที่สานยึดกันแน่นจะมีรูปเป็นผลึกสี่เหลี่ยมผืนผ้า (crystalline regions) ส่วนที่สานกันหลวมๆ นั้นเป็นบริเวณที่มีรูปร่างไม่แน่นอน (amorphous regions) ฉะนั้นใน cellulose แต่ละ fibril จะมีหลายผลึกสี่เหลี่ยมผืนผ้าและหลายส่วนที่รูปร่างไม่แน่นอน และในส่วนที่เป็นผลึกนี้จะทนทานต่อการทำลายของเอนไซม์ ทำให้เอนไซม์แทรกซึมเข้าไปในระหว่าง fibril ที่สานกันไม่ได้ ผนังเซลพืชจะมีสาร pectin, lignin หรือ hemicellulose ปริมาณต่างๆ กัน เชื่อมอยู่ตลอด
เอนไซม์ย่อย cellulose ที่เชื้อสร้างขึ้นเพื่อทำลายผนังเซลและ microfibril ทำให้อ่อนตัวเพื่อสะดวกต่อการแทงผ่านและเป็นอาหารของเชื้อ หากเกิดในกลุ่มท่อลำเลียง Cellulose ที่สลายตัวและมีโมเลกุลใหญ่จะเป็นอุปสรรคต่อการเคลื่อนย้ายน้ำในพืช เชื้อสาเหตุโรคที่สร้างเอนไซม์ย่อย cellulose ได้ มีหลายชนิดที่เป็นพวกบักเตรี เชื้อรา ไส้เดือนฝอย พืชชั้นสูงที่เป็นปรสิต และรา saprophyte บางชนิดที่อยู่ใน class Basidiomycetes
การย่อย cellulose ของเอนไซม์ มีกลไกที่เกี่ยวข้องในปฏิกริยาอยู่ 2 ทฤษฎี
1. ปฏิกริยาเกิดจากเอนไซม์ชนิดเดียว (unienzyme theory) การย่อย cellulose ไปเป็น glucose เกิดขึ้นโดยเอนไซม์ cellulase ชนิดเดียว
2. ปฏิกริยาเกิดจากเอนไซม์อย่างน้อย 2 ชนิด (multienzyme theory) คือ C1 กับ Cx โดย C1 เป็นเอนไซม์ชนิดที่ละลายผลึกสี่เหลี่ยมผืนผ้าของ fibril ด้วยการไฮโดรไลซ์ หรือกลไกอื่นที่ยังไม่ทราบ
cellulose ธรรมชาติ เช่น ฝ้าย ไปเป็น cellulose ที่มีขนาดสั้นลง (shorter linear polyanhydro glucose chains) เพื่อเปิดทางให้เอนไซม์ Cx แทรกซึมถึงเพื่อย่อย cellulose เป็นโมเลกุลเล็กๆ ต่อไป เช่น cellobiose แล้วเอนไซม์ ß -glucosidase ย่อยไปเป็น glucose ในที่สุด เอนไซม์ C1 นั้นจะเกิดจากเชื้อสาเหตุโรคซึ่งย่อย cellulose ตามธรรมชาติได้เท่านั้น ส่วน Cx ย่อย cellulose ธรรมชาติไม่ได้ แต่จะย่อยglucosidic bond เป็นแบบสุ่มเลือก และปลายสุด เรียกว่า Cx endo หรือ Cx exoenzymes ตามลำดับ
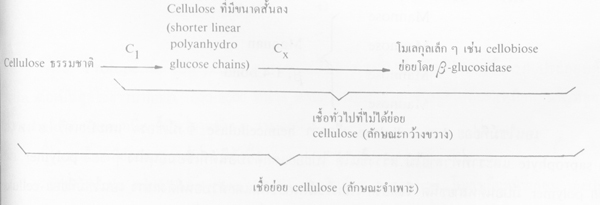
ภาพขั้นตอนการย่อยของ cellulose ธรรมชาติ (ที่มา: Goodman et al. 1967)
การย่อย cellulose จะยากหรือง่ายขึ้นอยู่กับลักษณะโครงสร้างของ cellulose fiber รวมทั้งปริมาณความชื้นของ fiber ขนาดและการแพร่กระจายของโมเลกุลเอนไซม์ที่เกี่ยวข้องกับขนาดและคุณสมบัติ ของช่องเล็กๆ ที่มีอยู่ระหว่าง microfibril ปริมาณของผลึก fibril การต่ออนูของ cellulose และธรรมชาติของสารที่อยู่ร่วมระหว่าง fibril
Hemicelluloses and hemicellulases
Hemicellulose ในช่องว่างระหว่าง microfibril ของผนังเซล มีสูตรทางเคมีที่ซับซ้อนด้วย polysaccharide ที่แตกต่างกัน เกิดจากการรวมกันของน้ำตาลที่เท่ากัน หรือโมเลกุลเล็กกว่า เช่น xylans, arbans, mannans และ galactans. Hemicellulose พบมากที่ผนังเซลของไม้ใหญ่และแก่เต็มที่ หญ้าและเมล็ดพืช Hemicellulose ในเนื้อเยื่อพืชชนิดต่างๆ มีปริมาณประมาณ 11-32% Hexose หรือ pentose ในผนังเซลที่เป็นส่วนประกอบของโมเลกุล hemicellulose มีความเข้มข้นแตกต่างกันไปคือ พบ gluecose มากที่สุด 55-73% xylose 9-39% galactose 1-17% mannose 0.4-16% arabinose น้อยกว่า 3.5% และมี rhamnose, methyl rhamnose, methyl ในปริมาณเพียงเล็กน้อย การเชื่อมกันของน้ำตาลเป็น henucellulose มีดังนี้

เอนไซม์ที่ย่อย hemicellulose เรียกว่า hemicellulase ซึ่งมีเชื้อรา และบักเตรี สาเหตุโรครา saprophyte และราที่ทำลายไม้ สร้างขึ้นได้ ไปย่อยแตกตัวบอนด์ที่เชื่อมอนุต่างๆ ของ polymer เนื่องจาก polymer มีบอนด์หลายชนิด จึงต้องการเอนไซม์หลายชนิดแตกตัวบอนด์ดังกล่าว เอนไซม์ที่ย่อย cellulose ของเชื้อราบางชนิดก็สามารถย่อยhemicellulose
Pectic substances and pectolytic enzymes
สารเพคตินเป็นส่วนประกอบของสารเชื่อมอยู่ระหว่างเซล เป็นชั้นใต้ cuticle ในเซล epidermis และในช่องว่าง cellulose fibril ในผนังเซลชั้นแรก (primary cell wall) สาร pectin ประกอบด้วยโปลิเßมอร์ของกรด galacturonic เชือมต่อกันที่ตำแหน่ง α11-4 และมักพบบ่อยว่ามี arabinose และ galactose เชื่อมอยู่ด้วยแต่ไม่ทราบตำแหน่ง สาร pectin มีลักษณะต่างกันไปที่ carboxyl ถูกแทนที่ด้วย methyl group. Polymer ของกรด galacturonic แบ่งออกได้เป็น 3 กลุ่ม โดยอาศัยความยาวของลูกโซ่และระดับของการถูกแทนทีด้วย methyl group คือ
1. Pectic acid เป็น polymer ของ d-galacturonic acid ที่เชื่อมต่อกันที่ตำแหน่ง α ,1-4 ไปยาวประมาณ 100 หน่วย มีคุณสมบัติละลายน้ำ และตกตะกอนได้ง่ายด้วยแคลเซียม และแมกนีเซียมไอออน เกิดเป็นเกลือ แคลเซียมและ magnesium pectate ที่ไม่ละลายนํ้า พบมากใน middle lamella
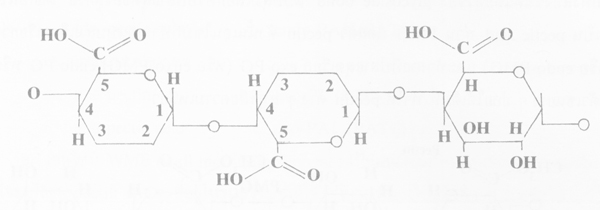
2. Pectin or pectinic acid เป็น methyl ester ของ pectic acid ที่มี polygalacturonic acid ยาวประมาณ 200 หน่วย ซึ่งถ้ากลุ่ม carboxyl ถูก esterified ด้วย methyl 75% หรือมากกว่า จะเป็น pectin หากน้อยกว่า จะเป็น pectinic acid
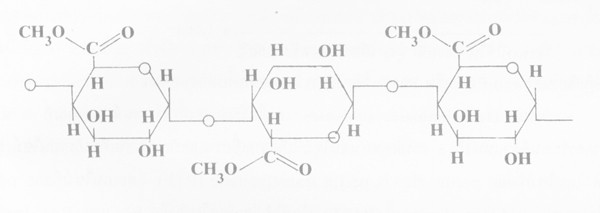
3. Protopectin เป็น pectin ธรรมชาติที่ไม่ละลายน้ำ มีอยู่ในผนังเซลชั้นแรก ประกอบด้วย pactin ต่อเป็นลูกโซ่ยาวมากกว่า 1000-2000 หน่วย และอาจพบเชื่อมข้ามกับลูกโซ่อื่นที่เหมือนกันหรือลูกโซ่ของ cellulose นอกจากนี้อาจมีนํ้าตาลต่างๆ ร่วมอยู่ด้วย
เอนไซม์ที่ย่อยสาร pectin มีดังนี้
1. Pectin methyl esterase (PME) เอนไซม์จะ hydrolyze โดยเคลื่อนย้ายกลุ่ม methyl จาก pectin หรือ pectinic acid ให้ pectinic acid หรือ pectic acid ตามลำดับและเมธิลแอลกอฮอล PME พบทั่วไปในพืชชั้นสูง และมีน้อยกว่ามากในจุลินทรีย์ต่างๆ
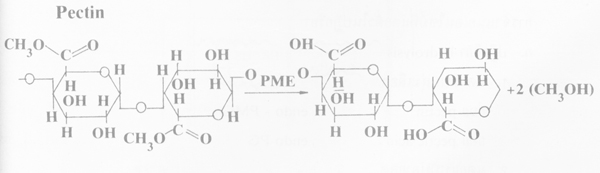
2. เอนไซม์ที่แตกตัวในปฏิกริยา (chain splitting enzymes) เอนไซม์จะไปแตก glycoside bond ที่ตำแหน่ง α ,1-4 ให้ d-galacturonic acid ซึ่งมีเอนไซม์ที่เกี่ยวข้องอยู่ 2 แบบคือ
ก. Glycosidases ได้แก่ polygalacturonase (PG) polymethyl galacturonase (PMG) เอนไซม์ทั้งสองชนิดแตกตัวของ glycoside bond แบบสุ่มเลือกหรือปลายสุดของลูกโซ่ โดยเอนไซม์ PG จะแตกเป็น pectic acid ส่วน PMG แตกตัว pectin ดังนั้นเอนไซม์ที่ทำลายแบบสุ่มเลือกเรียกว่า endo PG (หรือ endo-PMG) และทำลายที่ปลายสุดเรียก exo-PG (หรือ enxo-PMG) endo PG หรือ PMG จะแตกตัวเป็นยาวๆ ย่อยได้เต็มที่ ทำให้ pectin ต่างๆ สูญเสียความเหนียว

ส่วนปฏิกริยาของ exo-PG หรือ PMG ได้จากการวัดน้ำตาลที่ปล่อยออกมา
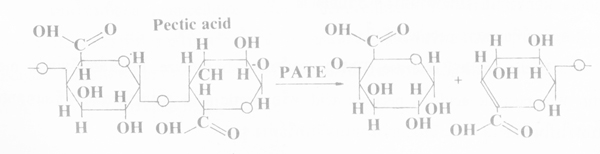
ข. Transeliminases or lyases เอนไซม์แตกตัวที่คาร์บอนด์ตำแหน่งที่ 4 และเอา H ออกจากคาร์บอนด์ตำแหน่งที่ 5 การย่อยของเอนไซม์นี้เกิดจากเชื้อราและบักเตรีเช่นเดียวกับเอนไซม์ hydrolyze
pectin เอนไซม์ที่ย่อย pectin เรียกว่า pectin transeliminase (PTE) และเอนไซม์ที่ย่อย pectic acid เรียกว่า pectic acid transeliminase (PATE) ชื่อที่ใช้เรียกเอนไซม์นี้คือ PL และ PAL (pectin lyase
and pectic acid lyase) ตามลำดับ โดยใช้คำ lyase แทน transeliminase ปฏิกริยาของเอนไซม์มีทั้งแบบสุ่มเลือกและที่ปลายสุด
การจำแนกเอนไซม์ที่แตกตัวในปฏิกริยา
ก. ปฏิกริยา hydrolysis
1. แตกแบบสุ่มเลือก
ย่อย pectin endo – PMG
ย่อย pectic acid endo-PG
2. แตกแบบปลายสุด
ย่อย pectin exo-PMG
ย่อย pectic acid exo-PG
ข. ปฏิกริยาแบบเคลื่อนย้ายและกำจัดแขนเชื่อม
1. แตกแบบสุ่ม
ย่อย pectin endo PL (PTE)
ย่อย pectic acid endo PAL (PATE)
2. แตกแบบปลายสุด
ย่อย pectin exo-PL (PTE)
ย่อย pectic acid exo-PAL (PATE)
3. เอนไซม์ WME (wall modifying enzyme) เป็นเอนไซม์ที่พบใหม่จาก Aspergillus niger เอนไซม์นี้จะเปลี่ยน pectin ต่างๆ ที่อยู่ในรูปธรรมชาติให้อยู่ในรูปที่เอนไซม์ตัวอื่นๆ ย่อยต่อไป
4. Protopectinase เป็นเอนไซม์รวมๆ ที่เตรียมเพื่อการย่อย protopectin ให้เป็น pectin ที่สั้นลง เอนไซม์นี้เป็นแบบ PG หรือ PMG ทำงานร่วมกับ PME
โปรตีนและเอนไซม์ย่อยโปรตีน (Proteins and proteolytic enzymes)
โปรตีนในผนังเซลพืชมี hydroxy proline เป็นส่วนประกอบอยู่มาก โปรตีนเกี่ยวข้องกับการเจริญเติบโตของผนังเซลที่เป็นตัวกลางเชื่อมระหว่างเส้น microfibril ของ cellulose (cross-linkages) โปรตีน ของผนังเซลนี้เรียกว่า extensin ซึ่งมีส่วนประกอบของ galacto-araban เชื่อมระหว่างโปรตีนกับ cellulose อยู่ด้วย (protein-galacto-araban) การติดเชื่อมระหว่าง microfibril ของ cellulose นี้อาจมี disulfide เชื่อมร่วมด้วย (S-S bridges) โปรตีนนอกจากอยู่ในผนังเซลแล้ว ยังเป็นสารอยู่ในเซล
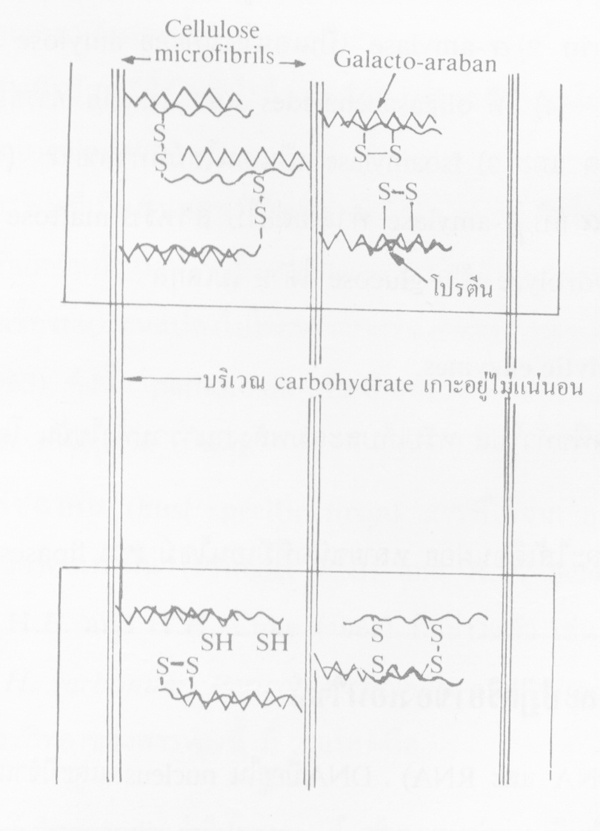
ภาพการเรียงตัวของ extension ในผนังเซลชั้นนอก (ที่มา:Goodman et al., 1967)
เอนไซม์ย่อยโปรตีนเป็น catalyst ของปฏิกริยา hydrolyze พวก peptide ที่เชื่อมเกาะกันเป็นโปรตีน แบ่งเอนไซม์โดยอาศัยขนาดของโมเลกุลได้ 2 ชนิด
1) Proteinases เป็นเอนไซม์ที่ hydrolyz โปรตีนไปเป็น polypeptides และ
2) Peptidases เป็นเอนไซม์ hydrolyze ขั้นต่อไป (polypeptides) ไปเป็น peptide และได้ amino acid ในที่สุด ความแตกต่างของเอนไซม์ทั้งสองยังไม่ทราบ เพราะพบว่า protinase สามารถย่อย peptide บางชนิด และ peptidase บางชนิดก็สามารถ hydrolyze โปรตีน Peptidase ที่ย่อย polypeptide ที่ N ของ amino acid ของ amino acid ปลายสุด เรียกว่า amino-peptidases และ peptidase ที่ย่อย peptide และโปรตีนที่ C ของ amino acid ปลายสุด เรียก carboxypeptidases สำหรับ amino acid ที่ได้จากการย่อยโปรตีนนี้ เชื้อจะดูดซึมไปใช้เพื่อสร้างโปรตีนของเชื้อเองต่อไปได้
แป้งและเอนไซมย่อยแป้ง (Starch and starch degrading enzymes)
แป้งเป็น polysaccharide ที่เซลพืชเก็บสำรอง แป้งได้รับจากการสังเคราะห์ใน chloroplast และ amyloplasts แป้งเป็น glucose polymers มีอยู่ 2 แบบคือ amylose และ amylopectin
Amylose เป็นโมเลกุล glucose ที่เชื่อมต่อกันที่ α- (1-4) ประมาณ 50-500 หน่วย มีน้ำหนักโมเลกุล 10,000-100,000 ส่วน amylopectin เป็นโมเลกุล glucose ที่เชื่อมต่อกันที่ α (1-6) เป็นหลักและแตกกิ่งด้วยการเชื่อมของโมเลกุลดังกล่าว Amylopectin มีจำนวนโมเลกุลประมาณ 250-5,000 หน่วย และน้ำหนักโมเลกุล 50,000-1,000,000
เชื้อส่วนมากจะย่อยแป้งและ polysaccharide ที่เก็บสำรองอื่นๆของพืช โดยมีเอนไซม์เกี่ยวข้องการ metabolism อยู่ 3 แบบ คือ
1) ß -amylase เป็นเอนไซม์ที่ย่อย amylose และ amylopectin ที่ α – (1 – 4) เป็น maltose เอนไซม์นี้สามารถย่อยได้สมบูรณ์ และยังสามารถย่อย amylopectin ที่เกาะอยู่นอก chain ได้เป็น dextrin
2) α -amylase เป็นเอนไซม์ที่ย่อย amylose และ amylopectin โดย hydrolyze สุ่มเลือกที่ α – (1 – 4) ให้ oligasaccharides และ dextrin การย่อยต่อไปก็จะได้ maltose, glucose และ dextrin ในที่สุด และ
3) Isoamylase เป็นเอนไซม์ที่ทำลาย α – (1 – 6) ของ amylopectin ให้ amylose ที่มีเชนสั้น ซึ่ง α กับ ß-amylase ทำลายต่อไป สำหรับ maltose นั้นจะแตกตัวโดยเอนไซม์ maltase ของจุลินทรีย์ ไป hydrolyze เป็น glucose ได้ 2 โมเลกุล
Lipids and lipolytic enzymes
Lipid มีอยู่ในเซลพืชทั่วไปสำหรับเก็บสะสมพลังงาน รวมทั้งไขมัน โดยเฉพาะในเซล epidermis และในเมล็ด
เชื้อรา บักเตรี และไส้เดือนฝอย หลายชนิดที่มีเอนไซม์ เช่น lipases และ phospholipidese ย่อยได้กรดไขมันต่างๆ
Nucleic acid และปฏิกริยาของเอนไซม์
Nucleic acid (DNA และ RNA), DNA มีอยู่ใน nucleus และมีจำนวนเล็กน้อยใน chloroplast, mitochondria ส่วน RNA มีอยู่ในทุกส่วนของเซล ใน cytoplasm,ribosome, mitochondria, chloroplast และ nucleolus โดย DNA & RNA เกี่ยวข้องในการถ่ายทอดพันธุกรรม การสังเคราะห์โปรตีน
Enzyme ที่ย่อย nucleic acid จากเชื้อราและบักเตรีมี ribonuclease (RNase) และ deoxy- ribonuclease (DNase) เป็นเอนไซม์ย่อย RNA และ DNA โดยการ hydrolysis ตามลำดับ ได้ mono- nucleotide ที่ประกอบด้วยโมเลกุลของเบส น้ำตาล และ ฟอสเฟต แล้วเอนไซม์ phosphatases จะ hydrolyze ต่อเป็นสารประกอบอนินทรีย์ฟอสเฟต และ nucleosides โดย nucleoside และ nucleotide นั้น อาจได้รับปฏิกริยาต่อโดยเคลื่อนย้ายกลุ่ม amino ออกไปจากเบสได้
สารพิษ (Toxins)
สารพิษ เป็นสารทุกชนิดที่เชื้อโรคสร้างและสามารถทำให้พืชเป็นโรคได้ ในอาการเดียวกับอาการโรคติดเชื้อที่เกิดตามธรรมชาติ สารนั้นอาจมีน้ำหนักโมเลกุลน้อยหรือมาก แต่สารที่มีน้ำหนักโมเลกุลตํ่าจะสามารถเคลื่อนไปถึงเซลของพืชภายในได้ง่าย
สารพิษนี้จะแตกต่างไปจากเอนไซม์ที่ไม่สามารถทำลายโครงสร้างของเนื้อเยื่อได้เหมือนเอนไซม์ แต่สารพิษไปมีปฏิกริยาต่อการ metabolism ใน protoplasm ของเซลโดยตรง
คำที่ใช้เกี่ยวกับสารพิษในเอกสารต่างๆ ทางโรคพืชมีดังนี้
Phytotoxin สารที่จุลินทรีย์สร้างขึ้น มีพิษไม่เฉพาะเจาะจงพืช ทำให้พืชมีอาการโรคเพียงเล็กน้อยหรือไม่มีเลย การสร้างสารพิษของเชื้อไม่มีความสัมพันธ์กับความสามารถในการทำให้พืชเป็นโรค เช่น lycomarasmin และ alternaric acid
Vivotoxin สารที่เกิดในพืชที่เป็นโรคติดเชื้อ ซึ่งเชื้อและ/หรือพืชอาศัยสร้างขึ้น ทำให้พืชเป็นโรคแต่ไม่ใช่สาเหตุเริ่มต้น เป็นเพียงส่วนหนึ่งที่ทำให้เกิดอาการโรค มีคุณสมบัติดังนี้
1. สารพิษนี้ต้องแยกได้จากพืชที่เป็นโรค แต่ไม่มีในพืชปกติ
2. สารพิษนี้มีคุณสมบัติทางเคมี
3. เมื่อนำสารพิษที่บริสุทธิ์นี้ฉีดเข้าไปในพืชปกติ พืชจะมีอาการโรคหรือมีอาการส่วนหนึ่งเกิดขึ้น เช่น fusaric acid และ piricularin
Pathotoxin สารพิษที่เป็นสาเหตุของโรคอย่างแท้จริง ทำให้เกิดโรคในพืชที่เป็นโรคง่าย เกี่ยวข้องกับลักษณะอาการของโรคที่เกิดขึ้นทั้งหมด เชื้อสาเหตุและสารพิษนี้จะแสดงอาการโรคแก่พืชอาศัยเฉพาะ เจาะจง และความรุนแรงของเชื้อจะเปลี่ยนแปลงไปโดยตรงกับความสามารถในการสร้างสารพิษนี้ (Wheeler, H.E. and H.H. Luke, 1963) ดังนั้น pathotoxin เป็นเชื้อที่เป็นสารเคมี (chemopathogens) ทำให้เกิดโรคได้เหมือนเชื้อสาเหตุ เช่น victorin เป็นต้น
สารพิษเฉพาะพืชอาศัย (host specific toxin) สารที่ได้จาก metabolism ของจุลินทรีย์ สาเหตุโรคที่มีพิษเฉพาะกับพืชอาศัยเท่านั้น (Pringle, R.B. and R.P. Scheffor, 1964) ซึ่งตรงกับ pathotoxin ของ Wheeler, H.E. and H.H. Luke (1963) ที่กล่าวแล้ว เช่น victorin (เกิดจาก Helminthosporium victoriae, H. carbonum, Periconia circinata and Alternaria kekuchiana etc.) ดังรายละเอียดปฏิกริยาทางสรีระวิทยาของสารพิษนี้ มี 2 แบบ คือ
ก. สารพิษทำให้เกิดอาการแก่พืชมาก หรือทั้งหมด
Victorin เกี่ยวข้องกับการเจริญของโรค ดังนี้
1. เฉพาะ strain ของเชื้อราที่สร้างสารพิษได้เท่านั้นที่ทำให้เกิดโรค
2. พืชที่เป็นโรคง่ายต่อเชื้อนี้จะเป็นโรคง่ายต่อสารพิษนี้ พืชที่ต้านทานต่อโรคจะต้านทานต่อปฏิกริยาของสารพิษด้วย และพืชที่มีความต้านทานต่อโรคปานกลางก็จะมีปฏิกริยาต่อสารพิษปานกลางเช่นกัน
3. สารพิษจะเป็นสาเหตุให้มีการเปลี่ยนแปลงทางชีวเคมีแก่พืชได้เหมือนกับเชื้อสาเหตุโรคของพืชนั้น
4. สารพิษที่เกิดจากสปอร์ขณะงอกที่จุดสัมผัส พืชในการแทงผ่านพืชอาศัยของเชื้อราสาเหตุสารพิษนี้หากนำไปหยดที่จุดสัมผัสของเชื้อที่ไม่ใช่สาเหตุบนโรคพืชนั้น ก็จะทำให้พืชเกิดโรคได้เช่นเดียวกัน
5. สารพิษนี้แม้เพียงเล็กน้อยก็สามารถมีผลต่อกิจกรรมทางชีวเคมีของพืชได้สูงมาก
ปฏิกริยาของสารพิษ victorin จะไปทำลายความสามารถของเยื่อหุ้มเซลพืชอาศัย ในการยอมให้ของเหลวผ่านเข้าออก และอัตราการหายใจของพืชเพิ่มหรือลดลง การใช้ฟอสเฟตและ amino acid เปลี่ยนไป ลดกิจกรรมเอนไซม์ amylase ที่ชั้น aleurone ของเมล็ด เพิ่มกิจกรรมการใช้คาร์บอนด์ไดอ๊อกไซด์ของพืชคือ
Periconia toxin สารที่มีพิษมากที่สุดที่เกิดจากเชื้อ ไม่เฉพาะเจาะจง แต่สารพิษที่เกิดจากเชื้อ Periconia circinata สาเหตุโรคไมโลของข้าวฟ่าง (milo disease of sorghum) มีคุณสมบัติเฉพาะเจาะจงสูงคล้ายสารวิคตอริน
สารนี้จะยับยั้งการเจริญของรากข้าวฟ่างเฉพาะพันธุ์ข้าวฟ่างที่เป็นโรคง่าย ไม่มีผลต่อพันธุ์ต้านทานโรค เป็นสารประกอบเคมีพวกเดียวกับวิคตอริน คือเป็นอนุพันธุ์ของโปลีเพพตายด์ อาการของโรคที่เกิดจากสารพิษนี้ จะเหมือนกับที่เกิดจากเชื้อ
Colletotin สารพิษที่เกิดจากเชื้อ Colletotrichum fuscum สาเหตุโรคของ foxglove (Di. gitalis spp) ที่ลำต้นโดยบริเวณที่เป็นโรคจะมีอาการชุ่มนํ้าในระยะแรก แล้วจะบุ๋ม และมีอาการ necrosis ในที่สุด สำหรับที่ใบแผลจะชุ่มน้ำเป็นจุดแล้วเปลี่ยนไปเป็น necrosis การเกิดชุ่มน้ำที่แผลในระยะแรกเนื่องจากการเคลื่อนของน้ำในเซลออกสู่ช่องว่างระหว่างเซลสารพิษนี้ไปเปลี่ยนความสามารถในการยอมให้ของเหลวผ่านเข้าออก พืชที่มีความต้านทานต่อเชื้อจะมีความต้านทานต่อสารพิษ สำหรับมันฝรั่งที่มีภูมิต้านทานต่อการติดเชื้อนี้อาจมีอาการของโรคที่เกิดจากสารพิษนี้ได้บ้าง
Wildfire toxin สารพิษที่เกิดจากเชื้อ Pseudomonas tabaci สาเหตุโรคไหม้ลามทุ่งของยาสูบ มีอาการเป็นจุดสีเหลือง ตรงกลางชุ่มน้ำ บนใบทั่วไป จุดขยายใหญ่ขึ้นและตรงกลางเปลี่ยนเป็นสีน้ำตาลเนื่องจากเนื้อเยื่อพืชตาย และเป็นวงรอบสีเขียวซีด แผลอาจลามติดต่อกันทำให้แผลใหญ่ขึ้น สารพิษที่เชื้อสร้างขึ้นจะทำลายเซลก่อนเชื้อเข้าสู่พืช แยกเชื้อบักเตรีสาเหตุโรคได้เฉพาะจากบริเวณกลางแผล สารพิษที่เชื้อสร้างขึ้นในอาหารเลี้ยงเชื้อ สามารถทำให้พืชเป็นโรคได้ทั้งในยาสูบที่เป็นโรคง่ายหรือยากก็ตาม จึงเป็นสารพิษที่ไม่เฉพาะเจาะจงพืช
สารพิษนี้ไปรบกวนการ metabolism ของ methionine ซึ่งเป็น amino acid ที่จำเป็น และทำให้เกิดวงรอบสีเขียวซีดรอบแผล
ข. สารพิษทำให้เกิดอาการแก่พืชบางส่วน
Piricularin สารพิษกึ่งเฉพาะเจาะจงพืช (semispecific) เกิดจากเชื้อ Piricutaria oryzae สาเหตุโรคใบไหม้ของข้าว โดยพันธุ์ที่เป็นโรคง่ายจะได้รับความเสียหายจากสารพิษนี้มากกว่าพันธุ์ที่ต้านทานโรค
Piricularin มีพิษกับพืชชั้นสูงและจุลินทรีย์อื่นๆ เชื้อราสามารถสร้างสารพิษนี้ในอาหารเลี้ยงเชื้อได้ในปริมาณมาก และพบว่าสารพิษนี้เสื่อมฤทธิ์ได้ด้วยโปรตีน (piricularin binding protein) และโปรตีนทำลายคุณสมบัติของสารพิษในการยับยั้งเชื้อรา แต่ไม่สามารถเปลี่ยนแปลงในการเป็นพิษของสารที่มีต่อต้นข้าว รายละเอียดต่างๆ ของโปรตีนนี้ในพืชที่เป็นโรคยังไม่ทราบ
Fusarial wilt toxin สารพิษเกิดจากเชื้อ Fusarium ลักษณะอาการของโรคเหี่ยวนี้เกิดจากการเจริญของผิวใบด้านบนเร็วมากกว่าปกติ ทำให้ใบห้อยลง (epinasty) การอุดตันและเป็นสีน้ำตาลของ xylem, necrosis และการเหี่ยวอาจเป็นปฏิกริยาของ vivotoxin ร่วมกันหลายชนิด โดยแต่ละชนิดของสารพิษทำให้เกิดเพียงส่วนใดส่วนหนึ่งของอาการทั้งหมด ไม่เฉพาะเจาะจงพืช และเป็นสาเหตุทำให้พืชมีการเปลี่ยนแปลงทางสรีระ เช่น fusaric acid เกิดจากราหลายสกุล และหลายชนิด โรคเหี่ยวของมะเขือเทศที่เกิดจาก Fusarium oxysporum lycopersici มี fusaric acid ที่เชื้อสร้างขึ้นร่วมอยู่ด้วย โดยไปลดกิจกรรมต่างๆ ของระบบหายใจของเนื้อเยื่อที่ลำต้น ซึ่งปกติตัวเชื้อเองนั้นไปทำให้มีการหายใจเพิ่มขึ้น Nishimura, S. (1962) กล่าวว่าโรคเหี่ยวนี้มีสารพิษร่วมด้วย 3 ตัว คือ fusaric acid, phytonivein และเอนไซม์ย่อย pectin
ปฏิกริยาของ fusaric acid ในพืชจะเกี่ยวกับกิจกรรมของการยอมให้ของเหลวผ่านเข้าออกเซล และแตกการสมดุลย์ของไอออนสารอนินทรีย์ และของ amino acid ไปยับยั้งการทำงานของเอนไซม์ polyphenoloxidase และตกตะกอนธาตุเหล็ก เป็นสาเหตุให้ขาดธาตุเหล็ก
ค. สารพิษทำให้เกิดอาการต่างๆ เพียงเล็กน้อย
Lycomarasmin และ alternaric acid เป็น phytotoxin ที่ไม่เฉพาะเจาะจงพืชทำให้พืชมีอาการเพียงเล็กน้อยหรือไม่มีเลย เกิดจากเชื้อ Fusarium oxysporum and Alternaria solani การเกิดสารพิษและการเกิดโรคของเชื้อรานี้ไม่มีความสัมพันธ์กัน
Lycomarasmin ทำให้มะเขือเทศมีใบงอขึ้นแล้วเนื้อเยื่อระหว่างเส้นใบเกิดนิโครซีส
Alternaric acid ทำให้เกิดแผลเป็นหย่อมบนใบและลำต้นของมะเขือเทศ โดยเป็นจุดสีน้ำตาลถึงสีดำ มีลักษณะเป็นวงแหวนขอบไม่เรียบ สีเข้มอยู่ตรงกลาง การเกิด chlorosis และใบร่วงเป็นลักษณะประจำของสารพิษนี้ต่อพืช
สารควบคุมการเจริญเติบโต (Growth regulators)
สารควบคุมการเจริญเติบโตของพืช เป็นสารประกอบที่มีปฏิกริยาเป็นโฮรโมนที่สำคัญมากที่สุด ได้แก่ auxins, gibberellins, cytokinins, เอธลีน และสารยับยั้งการเจริญต่างๆ (growth inhibitors) สารดังกล่าวทำหน้าที่ควบคุมการเจริญเติบโต พืชต้องการเพื่อขยายขนาดเซลตามกว้างและยาว กระตุ้นการทำงานของเซล การพักตัว และกิจกรรมต่างๆ ในบางกรณีก็สนับสนุนการสังเคราะห์ m RNA ให้เกิดเอนไซม์ที่มีคุณสมบัติเฉพาะ ควบคุมปฏิกริยาทางชีวะเคมีและสรีระวิทยาของพืช
สารควบคุมการเจริญเติบโตที่เชื้อสร้างขึ้นนั้นจะมีปฏิกริยาคล้ายกับสารที่พืชปกติสร้าง เมื่อเชื้อสร้างสารเหล่านี้ขณะพืชเกิดโรค ก็ทำให้พืชมีปฏิกริยาต่างๆ เนื่องจากปริมาณของสาร ตำแหน่งที่พืชได้รับและเวลาที่พืชต้องการ ไม่สอดคล้องกับความต้องการเพื่อการเจริญของพืชตามปกติ และเชื้ออาจสร้างสารที่แตกต่างไปจากพืชปกติต้องการอีกด้วย
สารที่เชื้อสร้างและแตกต่างไปจากสารที่พืชปกติสร้างนี้ จะทำให้การเจริญของพืชเปลี่ยนแปลงผิดปกติไป ตลอดจนเชื้ออาจสร้างสารที่ไม่ใช่โฮรโมน แต่อาจไปกระตุ้นระบบการสร้างโฮรโมนของพืชให้สร้างโฮรโมนมีจำนวนเกินความต้องการของพืชปกติ เป็นสาเหตุให้พืชมีการเจริญผิดปกติไป และในทางตรงกันข้าม เชื้ออาจสร้างสารที่ไปยับยั้งการทำงานของสารบางชนิดที่ควบคุมการเจริญของพืชตามปกติ หรือไปทำให้ระบบการสร้างโฮรโมนของพืชไม่ทำงาน
ดังนั้นจึงเชื่อได้ว่าเชื้อโรคเกี่ยวข้องกับกลไกปฏิกริยาต่างๆ ของพืช ทำให้เกิดการไม่สมดุลย์ของ ระบบโฮรโมนพืช มีผลให้การเจริญของพืชผิดไป เช่น เจริญมากกว่าปกติ แตกพุ่ม รากแตกฝอยมาก ลำต้นเปลี่ยนรูปแคระแกรน ใบร่วง ตาชงักงัน ใบห่อ เป็นต้น
Auxins
Auxins ที่อยู่ในพืชตามธรรมชาติเป็น IAA (ß-indole-3-acetic acid) เกิดในเนื้อเยื่อเจริญของใบอ่อน ปลายยอด มีอัตราเข้มข้นประมาณ 5 ไมโครกรัมต่อน้ำหนักพืช 1 กก. IAA เคลื่อนที่จากเนื้อเยื่ออ่อนสีเขียวไปยังเนื้อเยื่อที่แก่กว่าได้เร็ว แต่สลายตัวได้ด้วยเอนไซม์ IAA oxidase ซึ่งเป็น oxidative enzyme และอาจมี peroxidase enzyme ร่วมด้วย (oxidase -peroxidase) ดังสมการ

IAA เกิดขึ้นจากการเปลี่ยนแปลงมาจาก tryptophan และสารประกอบของ indole อื่นๆ ความสมดุลย์ IAA ในพืชควบคุมโดยเอนไซม์ และปฏิกริยายับยั้งของฟีนอล กับ catalase
IAA มีผลเกี่ยวข้องกับพืช ดังนี้
1. ส่งเสริมการเจริญของเซลพืชทางด้านยาว โดย pectins และ hemicelluloses ส่วนประกอบของเซลที่มี Ca2+ และ/หรือ Mg2+ เชื่อมอยู่ในรูปของเกลือ pectate ถูกเคลื่อนย้ายออกไปเป็น free carboxyl group ให้ pectic acid หรือ pectinic acid ทำให้ผนังเซลมีความยืดหยุ่น ขยายยาวออกไป
2. IAA ทำให้ความสามารถในการไหลผ่านของๆ เหลวเข้าออกเยื่อหุ้มเซลลดลง เนื่องจากเยื่อหุ้ม protoplast ของเซลดูดซับ IAA ไว้
3. IAA ไปควบคุมไม่ให้เกิดปฏิกริยาของเอนไซม์ PME ที่เยื่อหุ้มเซล เนื่องจาก IAA ที่เยื่อหุ้มเซลนั้นเกิดไปทำปฏิกริยารวมกับเอนไซม์ และ IAA ทำให้เซลขยายใหญ่ขึ้น

4. IAA อาจไปทำปฏิกริยากับสารประกอบฟีนอลต่างๆ เช่น catechol, chlorogenic acid และ caffeic acid ในขณะที่มีเอนไซม์ polyphenoloxidase อยู่ ทำให้สารประกอบฟีนอลเปลี่ยนไปเป็น
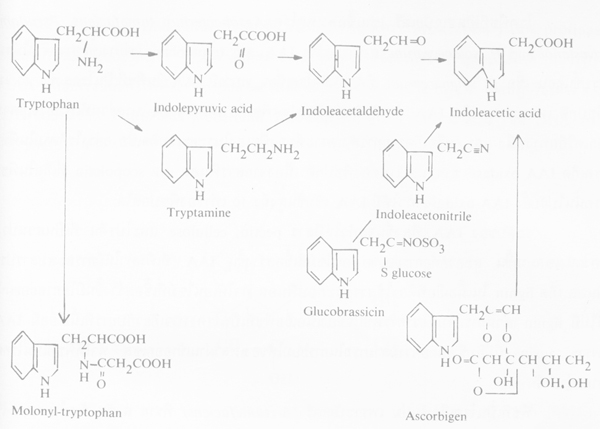
ภาพการสังเคราะห์ IAA จาก tryptophan และสารประกอบของ indole อื่นๆ (ที่มา: Goodman et al., 1967)
quinones ที่มี IAA เกาะร่วมอยู่ด้วย โดยเกาะจับที่ N ของ indole ring หรือเพิ่มที่ตำแหน่ง C ของ furane ring จึงทำให้ระดับของ IAA ในพืชมีน้อยลง ซึ่งการเพิ่มกิจกรรมของเอนไซม์ polyphenol oxidase ในพืชนี้เกิดขึ้นได้เมื่อพืชติดเชื้อ หรือพืชมีแผลเพราะเกิดจากวิธีกลต่างๆ
5. IAA ทำให้พืชมีอัตราการหายใจสูงขึ้น โดยไปกระตุ้นการทำงานของ oxidative enzymes เช่น dehydrogenases ตลอดจนระบบการเคลื่อนย้ายอีเล็คตรอนอื่นๆ
6. IAA อาจไปเปลี่ยนแปลงลักษณะการถ่ายทอดพันธุกรรมใน DNA ของเซล โดย IAAไปส่งเสริมการสังเคราะห์ m RNA และโปรตีน ซึ่งเกี่ยวข้องต่อการสังเคราะห์ RNA และเอนไซม์ ซึ่งการสังเคราะห์นี้มีหน่วยถ่ายพันธุ์ควบคุมอยู่
7. IAA มีปฏิกริยาร่วมกับ gibberellins และ cytokinins ดังนั้นระดับของ IAA ที่มีอยู่ในพืชไม่สมํ่าเสมอ อาจเกิดจากการสร้างมากไปหรือสลายตัวในระยะเวลานานไป จะเป็นสาเหตุทำให้พืชมีการเจริญเติบโตเปลี่ยนไปจากปกติ
พืชหลายชนิดที่เป็นโรคเกิดจากเชื้อรา บักเตรี และวิสา จะมีระดับ IAA สูงขึ้น แต่วิสาบางชนิด อาจทำให้ระดับ IAA ลดลงได้
โรคพืชที่เกิดจากเชื้อรา เช่น โรคใบไหม้ของมันฝรั่ง (Phytophthora infestans) โรคเขม่าดำของข้าวโพด (Ustilago maydis) โรคตายพรายของกล้วย (Fusarium oxysporum f. cubense) โรคไส้เดือนฝอยรากปม (Meloidogyne sp.) ฯลฯ นอกจากทำให้พืชที่เป็นโรคมี IAA เพิ่มสูงขึ้นแล้ว ตัวเชื้อเองก็สามารถสร้าง IAA ได้อีกด้วย อย่างไรก็ตาม IAA ที่เพิ่มขึ้นในพืชอาจเป็นจากเชื้อหรือพืช หรือทั้งสอง อย่างร่วมกัน และอาจทั้งหมดหรือบางส่วนเกิดจากการสลายตัวของ IAA เพราะ IAA oxidase ลดลง
โรคพืชที่เกิดจากบักเตรี เช่นเชื้อสาเหตุโรค Agrobacterium tumefaciens, Pseudomonas savastanoi and P. solanecearum สามารถสร้าง IAA จาก tryptophan แต่บางเชื้ออาจสร้างไม่สมบูรณ์ครบขั้นตอน เชื้อ P. solanacearum ที่ทำให้เกิดโรคเหี่ยว พบว่ามี IAA ในพืชที่เป็นโรคสูงถึง 10 เท่าของพืชปกติ เพราะว่าการเพิ่ม IAA นั้นเกิดติดต่อกันไปจนพืชเหี่ยวเต็มที่ IAA บางส่วนที่เพิ่มอาจเกิดจาทเชื้อบักเตรีอีกทางหนึ่ง และ/หรือเกิดจากการสังเคราะห์ของพืชเองในระยะเริ่มติดเชื้อ อย่างไรก็ตามในพืชที่เป็นโรคเกิด IAA oxidase ช้ากว่า 3 เท่าของพืชปกติ เนื่องจากสารฟีนอล เช่น scopoletin ที่เกิดในพืชที่เป็นโรคนั้นไปยับยั้ง IAA oxidase ทำให้มี IAA เข้มข้นสูงถึง 10 เท่าของพืชปกติได้
ระดับของ IAA ที่สูงขึ้นจะทำให้มีสาร pectin, cellulose และโปรตีน ที่เป็นส่วนประกอบของผนังเซลมากขึ้น สะดวกต่อการย่อยของเอนไซม์ที่เชื้อสร้างขึ้น IAA ที่เพิ่มทำให้มีการสะสมสารประกอบฟีนอล เกิด lignin ในเนื้อเยื่อที่ต้องใช้สารประกอบฟีนอล ทำให้เอนไซม์ที่เชื้อสร้างขึ้นมีโอกาสย่อยผนังเซลที่ไม่มี lignin ส่วนการเพิ่มอัตราการหายใจของเนื้อเยื่อพืชที่เป็นโรค อาจเกี่ยวกับการที่เนื้อเยื่อมี IAA สูงเพราะ auxin นี้มีอิทธิพลต่อความสามารถในการยอมให้ของเหลวผ่านเข้าออกเซล ทำให้พืชที่เป็นโรคคายน้ำมากกว่าปกติ
พืชที่เป็นโรคเกิดปุ่มปม เพราะบักเตรี (A. tumefaciens) ที่ราก ลำต้น ก้านใบ ฯลฯ เชื้อเข้าสู่พืชโดยทางแผล ทำให้มีการแบ่งเซลรอบแผลอย่างรวดเร็วเกิดขึ้นในระยะ 2-3 วันหลังการเกิดแผล เนื่องจากการกระตุ้นของเชื้อและปฏิกริยาโต้ตอบของเซลพืชต่อการทำลายของเชื้อ การกระตุ้นให้เกิดปุ่มปมนี้เรียกว่า TIP (tumor-inducing principle) การเจริญและแบ่งเซลให้เกิดปุ่มปมจะเกิดอย่างอิสระ ไม่เกี่ยวกับกิจกรรมของบักเตรี และอัตรการเติบโตและการแบ่งเซลนี้พืชควบคุมได้โดยใช้เวลาไม่มากนัก
เซลที่เป็นปุ่มปมของพืชจะมี IAA และ cytokinin สูงกว่าเซลปกติ แม้ว่าในเซลนั้นจะไม่มีเชื้ออยู่ภายในด้วยก็ตาม การเกิดปุ่มปมเป็นเพราะระดับของ IAA ที่มีอยู่ในเซลสูงกว่าระดับปกติ เนื่องจากมีการสังเคราะห์มากกว่าการสลายตัว แต่การที่เซลมี IAA สูงมากกว่าปกติอย่างเดียว จะไม่สามารถทำให้เซลเป็นปุ่มปมได้ สาเหตุที่เกี่ยวข้องต่อการเกิดนั้นยังไม่ทราบ แต่การเกิดเป็นปุ่มปมนั้นไม่เร็วขึ้นเมื่อถูกความร้อน นอกจากใช้หยดด้วย glutamine และ meso-inositol และจะเกิดปุ่มปมนั้นเร็วยิ่งขึ้นอีก เมื่อเพิ่ม aspartic acid, cytidylic acid และ guanalic acid สำหรับสารต่างๆ ดังกล่าวเกี่ยวข้องต่อการเกิดปุ่มปมอย่างไรนั้น ยังไม่ทราบ
เชื้อวิสาสาเหตุโรคส่วนมากทำให้พืชมีอาการแคระแกรน แตกตามากกว่าปกติ ลักษณะสัณฐานของส่วนต่างๆ ของพืชเจริญผิดปกติ อาการดังกล่าวคล้ายกับอาการของพืชที่เกิดจากความไม่สมดุลย์ของสารต่างๆ เพื่อการเจริญเติบโตของพืช พืชที่เป็นโรคส่วนมาก มี auxin ในระดับต่ำกว่าปกติ โดยเฉพาะในระยะที่พืชแสดงอาการเด่นชัด จึงทำให้พืชมีการเจริญเติบโตน้อย
Gibberellins
Gibberellin เป็นสารเคมีกลุ่มหนึ่งคล้าย IAA เป็นส่วนประกอบตามปกติในพืชสีเขียวและเป็นสารที่จุลินทรีย์หลายชนิดสร้างได้ Gibberellin ทำให้พืชเจริญแตกต่างกันไปตามชนิดของพืช Gibberellin เป็นโฮรโมนในคัพภะของเมล็ด กระตุ้นให้ขับเอนไซม์ เช่น -amylase ในชั้น aleurone ไปย่อยแป้งที่เก็บสะสมอยู่ และกระตุ้นเอนไซม์ในการแตกตัวของผนังเซลที่เกี่ยวกับการย่อย endosperm และทำให้เปลือกหุ้มเมล็ดอ่อนตัว นอกจากนี้ gibberellin ยังกระตุ้นหรือให้มีการสังเคราะห์เอนไซม์อื่นๆ เช่น protinase เพื่อไปย่อยโปรตีนใน endosperm ไปเป็น amino acid และ tryptophan ซึ่ง tryptophan นี้เป็นสารที่ให้กำเนิด IAA Gibberellin ทำให้พืชเจริญทางด้านยาวคล้ายกับสาเหตุที่เกิดจาก IAA โดยปฏิกริยาต่างๆ ที่เกิดขึ้นอาจคล้ายคลึงกัน ปฏิกริยาของ gibberellin ในเซลจะเกี่ยวข้องกับ nucleic acid เช่น DNA และ RNA ของระบบต่างๆ
ในพืชที่เป็นโรคโคนเน่าของข้าว ที่เกิดจากเชื้อรา Gibberella fujikuroi เชื้อสาเหตุโรคสร้าง gibberellin ทำให้พืชเจริญเติบโตอย่างรวดเร็วและมีอัตราสูงมากกว่าพืชปกติ
พืชที่เป็นโรคเกิดจากวิสา มีอาการและแคระแกรนเพราะเกี่ยวกับความเข้มข้นของ gibberellin ในพืชลดลง อาการของโรคจะชงักหายไปเมื่อใช้ gibberellin และอาการจะกลับคืนมาใหม่หากหยุดใช้
Cytokinins
Cytokinins เป็นปัจจัยที่สำคัญเพื่อาการเจริญเติบโตและให้เซลเจริญแตกต่างกัน Cytokinins ที่เกิดตามธรรมชาติ มีความเข้มข้นต่ำ พบในพืชสีเขียวที่เมล็ด และน้ำเลี้ยงของพืช Cytokinins เป็นอนุพันธ์ ของ adinine ซึ่งเป็นส่วนประกอบของ DNA และ RNA ได้แก่ kinetin (6-furfurylamino-purine), zeatin, IPA [N6 – Z2-ispentenyl) adenosine]
ปฏิกริยาของ cytokinins จะเกิดขึ้นได้ต้องมี IAA ร่วมอยู่ด้วย ทำให้เซลขยายใหญ่ และมีการแบ่งตัว หากมีสารใดสารหนึ่งเพียงอย่างเดียว ปฏิกริยานั้นก็ไม่เกิดขึ้น กลไกของการเกิดดังกล่าวยังไม่ทราบแต่
เข้าใจว่าจะคล้ายกับ gibberellin คือ เกี่ยวกับการสังเคราะห์ m RNA โดยการกระตุ้นของหน่วยถ่ายพันธุ์ เช่น โรคใบไหม้ของข้าวโอ๊ตที่เกิดจากเชื้อรา Helminthosporium สร้างสารพิษ victorin โรคจะรุนแรงขึ้นเมื่อมี cytokinin โดย cytokinin ไปเพิ่มปริมาณของสารพิษมากขึ้น ดังนั้น cytokinin อาจไปส่งเสริมเชื้อสร้าง pathotoxin และ cytokinin อาจมีส่วนเกี่ยวข้องต่อการเกิดอาการพุ่มแจ้ของพืชที่เกิดจากเชื้อรา และโรคไฟไหม้ลามทุ่งของยาสูบ ที่เกิดจากบักเตรี หากฉีดพ่น cytokinins บนยาสูบที่ได้รับสารพิษนี้ ยาสูบจะไม่มีอาการโรคเกิดขึ้น ส่วนโรคที่เกิดจากวิสา พบว่าหากใส่ kinetin ก่อนหรือหลังปลูกเชื้อแล้ว อาการโรคที่เกิดเป็นแห่งๆ จะมีจำนวนน้อยลง และพืชที่เป็นโรคแบบกระจายทั้งต้น อาการจะไม่รุนแรง เนื่องจากการทวีจำนวนของวิสา
ลดลง
สารยับยั้งการเจริญ (Growth inhibitors)
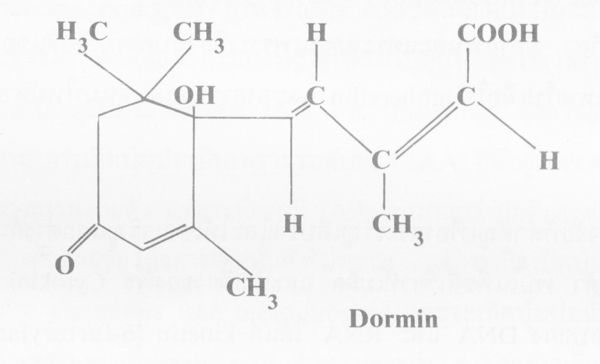
สารยับยั้งการเจริญเหล่านี้จะมีปฏิกริยาร่วมกับโฮรโมนและบางครั้งจะขัดขวางหน้าที่ของส่วนต่างๆ ในภายหลัง เช่น dormin หรือ abscisin II ทำให์ใบที่เพิ่งจะโผล่ออกจากตา ลอกเป็นสะเก็ด ซึ่งอาการ จะหายไปได้โดยการใช้ gibberellin หรืออีกนัยหนึ่ง dormin ไปยับยั้งการกระตุ้นของเอนไซม์ -amylase ใน endosperm ของเมล็ด ซึ่งอาจกล่าวได้ว่า dormin มีหน้าที่ต่อต้านสาร gibberellin ในพืช และในบางกรณีอาจไปยับยั้ง IAA ฉะนั้นหน้าที่ของสารยับยั้งนี้จะเกี่ยวร่วมกันอย่างใกล้ชิดกับความเข้มข้นของโฮรโมน หรือได้รับอิทธิพลจากโฮรโมนที่มีอยู่ในพืช และอาจเกิดขึ้นได้ในทางกลับกันด้วย
เอธลีน (Ethylene)
เอธลีนเกิดจาก amino acid, methionin ในเนื้อเยื่อพืช จะระเหยเป็นแก๊สผ่านทางใบสู่อากาศ สารประกอบเอธลีนในพืชที่เกิดจากเชื้อ จะทำให้ใบร่วงมีการหายใจเพิ่มขึ้น เช่น โรคของกล้วยที่เกิดจากPseudomonas solanacearum จะทำให้ผลแก่ก่อนกำหนด เนื่องจากการที่มีเอธลีนเข้มข้นสูงเพราะเชื้อสร้างขึ้น เป็นต้น
Polysaccharides
เชื้อรา บักเตรี และไส้เดือนฝอยศัตรูพืชบางชนิดปล่อยสารเป็นเมือก (mucilaginous substances) หุ้มผิวนอกไม่ให้ตัวเชื้อหรือไส้เดือนฝอยกระทบกับสิ่งแวดล้อมที่อยู่โดยรอบโดยตรง สารที่เป็นเมือกเหล่านี้ เป็น polysaccharide เป็นส่วนใหญ่ และมีจำนวนเล็กน้อยที่เป็น amino acid โปรตีน และ lipid ส่วนประกอบทางเคมีของ polysaccharide ก็แตกต่างกันไปอย่างกว้างขวางแล้วแต่ชนิดของจุลินทรีย์ เชื้อราและบักเตรีสร้าง polysaccharide ที่ประกอบด้วย glucose เป็นหลักและมี fructose, galactose, mannose, xylose, rhamnose, fucose และ glucoronic acid มีน้ำหนักโมเลกุลแตกต่างกันไปจากหลายร้อยถึงหลายล้านหน่วย
Polysaccharide เกี่ยวข้องกับโรคเหี่ยวของพืช โดยเชื้อราสาเหตุโรคทำลายกลุ่มท่อลำเลียงทำให้พืชเหี่ยว โดยทั่วไปบักเตรีสาเหตุโรคทุกสกุลสร้างได้ ปริมาณจะแตกต่างไปแล้วแต่ชนิด Polysaccharide นี้นอกจากเชื้อสร้างขึ้นโดยตรงแล้ว อาจเป็นปฏิกริยาของเชื้อไปแตกตัวสารต่างๆ ของพืชออกมา เชื้อบักเตรีสามารถสร้างได้ดีในปริมาณที่มากกว่าเชื้ออื่นๆ
ที่มา:ไพโรจน์ จ๋วงพานิช
