GENETICS OF PLANT-PATHOGEN INTERACTION
กลไกการเข้าทำลายของเชื้อและการป้องกันตนเองของพืชจะถูกกำหนด โดยพันธุกรรมของเชื้อและะพืชตามลำดับ การเปลี่ยนแปลงใดๆ ที่เกิดขึ้นทางพันธุกรรมของเชื้อจะทำให้ได้เชื้อ races ใหม่ๆ ซึ่งแต่ละ race จะมีความแตกต่างกันในความสามารถของการทำให้เกิดโรคและอาจเข้าทำลายพืชที่มีความต้านทานโรคได้ และในลักษณะเดียวกัน พันธุ์ใหม่ของพืชที่เกิดขึ้น อาจมีความต้านทานต่อโรคสูงขึ้น อันจะมีผลทำให้เชื้อ race ใหม่และพืชพันธุ์ใหม่นั้นมีแนวโน้มที่จะมีชีวิตยืนยาวออกไป โดยเชื้ออื่นๆ ที่ไม่สามารกทำให้พืชเป็นโรค และพืชพันธุ์อื่นซึ่งเป็นโรคง่ายกว่าก็จะสูญสิ้นไป การเกิดโรค และการเจริญของโรคจะดีต่อเมื่อเชื้อมี race ที่รุนแรงพบกับพืชที่เป็นโรคง่าย และโรคจะไม่เกิดขึ้นหากเชื้อไม่รุนแรงหรือพืชมีความต้านทานต่อโรค แต่ทั้งนี้ขึ้นอยู่กับสภาพแวดล้อม ซึ่งมีผลโดยตรงต่อเชื้อและ/หรือต่อพืช ปฏิกริยาที่มีต่อพืชและเชื้อโรค ขึ้นอยู่กับความสำเร็จในการต้านทานของพืชและความสามารถในการเป็นปรสิตของเชื้อภายใต้สภาพแวดล้อม ที่เอื้ออำนวยให้ วิทยาการทางพันธุศาสตร์ของการต้านโรคของพืช มีความสำคัญอย่างยิ่งต่อการผสมพันธุ์หาพันธุ์ที่ควบคุมโรคได้
กลไกนี้จะกล่าวเฉพาะการเปลี่ยนแปลงต่างๆ ที่เกิดกับเชื้อโรคเท่านั้น
การขยายพันธุ์แบบไม่ใช้เพศ (asexual reproduction)
การขยายพันธุ์แบบไม่ใช้เพศนี้ พบทั่วไปกับเชื้อรา มีการแบ่งเซลที่ nucleus ไม่รวมกัน เซลที่ได้ใหม่จะเจริญต่อไปอย่างอิสระและมีลักษณะของพันธุ์เหมือนเดิม แล้วเจริญเป็นเส้นใยที่ประกอบด้วยหลายเซล และเกิดสปอร์ที่ไม่แตกต่างไปจากพันธุกรรมเดิม แต่อย่างไรก็ตามกลไกการขยายพันธุ์แบบไม่ใช้เพศนี้ อาจมีการเปลี่ยนแปลงทางพันธุกรรมเกิดขึ้นได้ 2 ทางคือ
1. Mutation เป็นการเปลี่ยนแปลงในสารถ่ายทอดพันธุกรรมอย่างทันที ได้ลักษณะใหม่ที่ผิดไปจากพ่อแม่ Mutation เกิดขึ้นเองกับจุลินทรีย์ที่มีชีวิตต่างๆ ในธรรมชาติอย่างบังเอิญในระหว่างการแบ่งเซลเป็นผลจากการทวีจำนวนที่ไม่สมํ่าเสมอ หรือการจัดเรียงส่วนเล็กๆ ของสารถ่ายทอดพันธุกรรมของเซล การเกิด mutation จะมีอัตราสูงมากหากจุลินทรีย์อยู่ในสภาพที่ถูกแสงอุลตราไวโอเลท แสงรังสี X รังสีแกมม่า อุณหภูมิสูงหรือมีปฏิกริยากับสารเคมีบางชนิด เช่น epoxides, peroxide ฟีนอล alkaloid ต่างๆ เป็นต้น
Mutation ที่เกิดในระหว่างการแบ่งเซล mitosis ซึ่งเป็นการแบ่งของเซลตามปกติ โดยการแยกออกของ chromatin ของ nucleus ไปเป็นรูปคล้ายเส้นด้ายที่แบ่งออกเป็นส่วนๆ เรียกว่า chromosomes แต่ละ chromosome จะถูกแยกออกตามความยาวออกเป็นสองส่วน แต่ละส่วนจะไปอยู่ในแต่ละเซลที่เกิดใหม่ 2 เซล แล้วแต่ละส่วนที่อยู่ในเซลใหม่นั้น จะทวีจำนวนทันทีเพื่อเป็นเซลลูกที่มีหน่วยถ่ายพันธุ์เหมือนเซลแม่ อย่างไรก็ตามการทวีจำนวนของ chromosome นั้นในบางครั้งไม่ได้เกิดขึ้นทันที หรือในระหว่างการแยกออกของ chromosome ที่เป็นคู่อยู่นั้น มีเศษของ chromosome แตกหลุดออก ทำให้เซลลูกมีหน่วยถ่ายพันธุ์ที่มากกว่า และเซลลูกที่มีหน่วยถ่ายพันธุ์น้อยกว่าเซลแม่ เป็น 2 เซลแตกต่างกันที่ไม่เหมือนเซลแม่ การเปลี่ยนแปลงที่เกิดขึ้นนี้จะไปมีผลต่อลูกต่อไป
2. Heterokaryosis เป็นสภาพของเส้นใย หรือเซลของเส้นใยที่มี nucleus อยู่มากมาย ประกอบด้วยพันธุกรรมที่แตกต่างกัน Heterokaryosis เกิดจากการรวมกันของเส้นใยที่คล้ายกัน แต่มีพันธุกรรมแตกต่างกัน ซึ่งพบในราทั่วไป อาจทำให้มีการจัดกลุ่ม nucleus ที่มีชนิดแตกต่างกันใหม่ แต่ nucleus จะไม่รวมกัน
Heterokaryosis ไม่เหมือน dikaryosis ที่เส้นใยของ dikaryosis มี 2 nuclei และมีเพศตรงข้ามกัน (n +n) แต่ละ nucleus ก็จะแบ่งตัวส่งให้เซลที่เกิดใหม่
การขยายพันธุ์แบบใช้เพศ (Sexual reproduction)
การขยายพันธุ์ใช้เพศแบบ hybridization เป็นเหตุให้มีการจับคู่กันของหน่วยถ่ายพันธุ์ และแยกออกจากกันได้ลูกที่สืบมาจากพ่อแม่ที่มี race พันธุ์ (varieties) ชนิดหรือสกุลต่างกัน โดย hybridization ที่พบในราจะเริ่มจากการรวมกันของสอง nuclei ซึ่งแต่ละ nucleus มี chromosome n เดียว (haploid neuclei) Nucleus มี strain เป็น + และ – ต่างกัน หรือมีเพศตรงกันข้าม ทำให้ได้ nucleus มี chromosome 2n (diploid nucleus) คือ zygote cell แล้วจึงแบ่งตัวแบบ meiosis ต่อไปในระหว่างการแบ่งตัวแบบ meiosis นั้น chromosome ที่มีโครงสร้างลักษณะเหมือนกันของพ่อแม่จะจับคู่ขนานซึ่งกันและกัน แล้ว chromosome ที่จับคู่กันแล้วนั้นจะแบ่งเป็น chromatids การแลกเปลี่ยน chromatid ข้ามกันเพียงบางส่วน (crossing over) เกิดขึ้นได้ในการแลกเปลี่ยนคู่ของ chromosome การจับคู่กันใหม่ของสารถ่ายทอดพันธุกรรม จาก nucleus พ่อแม่จะเกิดใน zygote cell เมื่อแบ่งเซลแบบ meiosis แล้ว ทำให้ได้เซลที่แตกต่างซึ่งกันและกัน และต่างไปจากเซลพ่อแม่ Hybridization พบเกิดส่วนมากโดยทั่วไป และทำให้มีการเปลี่ยนแปลงในความสามารถของเชื้อต่อการเกิดโรคบ่อยๆ
เชื้อรา
สายพันธ์ของเชื้อราเห็นได้ชัด ในราสาเหตุโรคราสนิม โดย race ต่างๆ ของเชื้อราจะมีระดับความสามารถทำให้เกิดโรคแตกต่างกัน เนื่องจากเชื้อประกอบด้วยหน่วยถ่ายพันธุ์จำนวนหนึ่ง แต่ละหน่วยอาจควบคุมคุณสมบัติเฉพาะของระดับความสามารถในการทำให้เกิดโรค และพืชอาศัยเองก็มีหน่วยถ่ายพันธุ์ จำนวนหนึ่ง ที่แต่ละหน่วยอาจควบคุมความต้านทานโรค หรือการเป็นโรคง่ายของพืชอย่างใดอย่างหนึ่ง และจะมีหน่วยถ่ายพันธุ์เฉพาะของเชื้อที่จะมาทำลายหน่วยถ่ายพันธุ์ที่ต้านทานโรคของพืช ซึ่งจะทำให้เชื้อ race หนึ่ง เข้าทำลายพืชได้เฉพาะพืชอาศัยบางพันธุ์ (Gene for Gene concept; Florr, H.H. 1955)

ภาพไดอาแกรมแสดงการเปลี่ยนแปลงของเชื้อที่ขยายพันธุ์แบบใช้เพศ เนื่องจากการเเลกเปลี่ยนของ chromatid ข้ามกันเพียงบางส่วนระหว่างการแบ่งเซลแบบ meiosis แล้วเกิดการจับคู่กันใหม่ของสารถ่ายทอดพันธุกรรม ทำให้ได้เซลที่แตกต่างซึ่งกันและกันและต่างไปจากเซลพ่อแม่ (ที่มา : Agrios, 1969)
Puccinia graminis เชื้อราสาเหตุโรคราสนิมของธัญญพืช ซึ่งเป็นปรสิตถาวร เชื้อถูกแบ่งย่อยออกไปจาก species เนื่องจากเชื้อทำลายพืชเพียงเฉพาะพืชอาศัยเท่านั้น และพบว่ามีไม่น้อยกว่า 6 กลุ่มย่อย จึงให้ชื่อแสดงกลุ่มย่อยเป็น sub-species (พันธุ์ หรือชนิดย่อย) เช่น
P. graminis tritici ทำให้เกิดโรคกับข้าวสาลี
P. graminis hordei ทำให้เกิดโรคกับข้าวบาร์เลย์
P. graminis avenae ทำให้เกิดโรคกับข้าวโอ๊ต
P. graminis secalis ทำให้เกิดโรคกับข้าวไรน์
ซึ่งเป็นการเพิ่มชื่อลาตินคำที่สามต่อท้ายชื่อเดิม (binomial เป็น trinomial) แต่ละชนิดย่อยของเชื้อประกอบด้วย race จำนวนมาก เช่นพบว่า P. graminis tritici มีมากกว่า 240 races ชื่อประจำ race จะเขียนเป็นตัวเลขต่อท้าย ชื่อ sub-species
Race ที่เป็นเชื้อทำลายพืชเกิดในช่วงที่เชื้อมีการขยายพันธุ์แบบไม่ใช้เพศ เช่น race ของราสาเหตุโรคราสนิม เป็นระยะ uredospores สำหรับโรคเขม่าดำเชื้อที่ก่อโรคที่เป็น inoculum ตามธรรมชาติ เป็น chlamydospores, dikaryotic teliospore โดย nucleus จะรวมกันแล้วงอกและมีการแบ่งเซลแบบ meiosis ได้ basidiospore มี chromosome 1n เข้าทำลายพืช อย่างไรก็ตาม race ของราสาเหตุโรคเขม่าดำ ที่เข้าทำลายพืชยังใช้กลุ่มของ chlamydospore เป็นหลักอยู่เนื่องจากมีความสมํ่าเสมอในความสามารถของเชื้อในการทำให้เกิดโรค และกิจกรรมของเชื้อตลอดวงจรโรค ดังนั้น race ของเชื้อที่ทำลายพืชจึงมีอยู่ในช่วงขยายพันธุ์แบบใช้เพศ
บักเตรี
บักเตรีสาเหตุโรคพืชมีการเปลี่ยนแปลงเกิดขึ้นอย่างมากมาย แม้ว่าเชื้อบักเตรี จะมีการฃยายพันธุ์ เฉพาะแบบไม่ใช้เพศเพียงอย่างเดียว การเปลี่ยนแปลงอาจเป็นแบบชั่วคราว โดยเป็นการเปลี่ยนที่เกิดขึ้นทางสรีรวิทยา หรือทางสัณฐานวิทยา ซึ่งเกิดจากสภาพแวดล้อมที่เปลี่ยนไปทั้งทางฟิสิกส์หรือทางเคมีที่มีต่อบัทเตรี การเปลี่ยนแปลงชั่วคราวของเชื้อนี้อาจกลับคืนสู่สภาวะปกติได้ และการเปลี่ยนแปลงอีกแบบหนึ่งเป็นแบบถาวร เกิดจากการเปลี่ยนแปลงใน DNA ของบักเตรี การเปลี่ยนแปลงแบบนี้จะกลับคืนสู่ปกติได้ยาก มีความสำคัญต่อความสามารถของบักเตรีในการทำให้เกิดโรค
การเปลี่ยนแปลงที่เกิดกับบักเตรีสาเหตุโรค มีดังนี้
1. Mutation โดยบักเตรีจะแบ่งเซลจากหนึ่งเป็นสองขณะที่แบ่งเซล DNA ส่วนใดส่วนหนึ่ง อาจถูกเปลี่ยนแปลงไปเป็นผลให้เซลใหม่นั้นแตกต่างไปจากเซลเดิม การเกิด mutation นี้อาจเกิดขึ้นเองตามธรรมชาติ หรือสารเคมีต่างๆ หรือรังสี เช่นรังสีคอสมิค (cosmic rays) และอุลทราไวโอเลต
2. Transformation เกิดขึ้นได้กับบักเตรีสาเหตุโรคบางชนิดเท่านั้น เป็นวิธีง่ายที่สุดของการแลกเปลี่ยนสารถ่ายทอดพันธุกรรม โดยการสูญเสียของบักเตรีเซลหนึ่งด้วยการดูดซึมของ DNA ที่เหมือนกัน จากบักเตรีอีกเซลหนึ่ง
3. Conjugation โดยการจับคู่และสัมผัสกันของบักเตรีสองเซล แล้วส่วนน้อยของสารถ่ายทอดพันธุกรรมจากเซลหนึ่ง จะเคลื่อนย้ายเข้าไปเป็นสารถ่ายทอดพันธุกรรมของอีกเซลหนึ่งด้วย
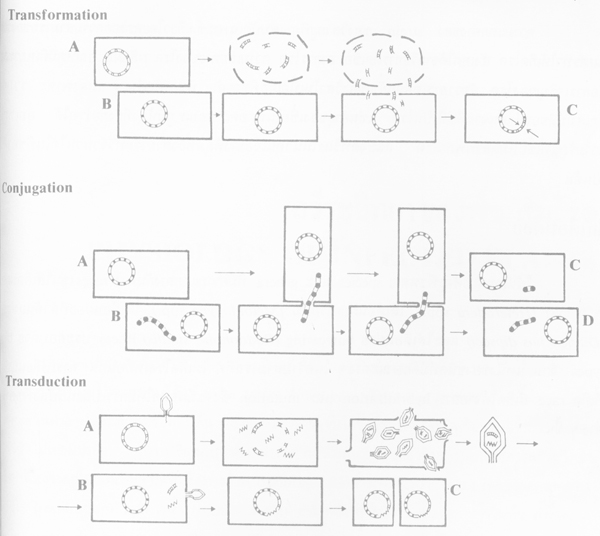
ภาพกลไกการเปลี่ยนแปลงพันธุกรรมของบักเตรี (ที่มา : Agrios, 1978)
4. Transduction โดย bacteriophage เข้าทำลายบักเตรี ทำให้ได้รับสารถ่ายทอดพันธุกรรมไว้ แล้วไปทำลายบักเตรีอีกเซลหนึ่ง จึงทำให้สารถ่ายทอดพันธุ์นี้ไปปะปนกับของบักเตรีเซลใหม่ ทำให้มีการเคลื่อนย้าย หรือมีการเปลี่ยนแปลงพันธุกรรมขึ้น
ความรุนแรงของเชื้อในการเกิดโรคแก่พืช อาจลดลงหากเป็นเชื้อที่ได้จากการเจริญบนอาหารเลี้ยงเชื้อ และเชื้อ strain ที่ไม่รุนแรง จะเจริญในอาหารเลี้ยงเชื้อได้ดีกว่าเชื้อ strain ที่รุนแรง
วิสา
การเปลี่ยนแปลงของวิสาสาเหตุโรคเป็นที่ยอมรับกันมาเป็นเวลานานแล้ว การถ่ายทอดวิสาสาเหตุโรคจากพืชที่เป็นโรคไปยังต้นอื่นๆ อาการโรคที่เกิดใหม่นั้นก็มักจะเหมือนต้นที่เป็นโรคอยู่เดิม เพราะในบางครั้งอนุภาคของวิสาบางส่วน อาจไม่เหมือนกับวิสาสาเหตุโรคจากต้นเดิม แต่อนุภาควิสาที่เหมือนเดิมมีมากกว่า จึงทำให้พืชที่เป็นโรคมีอาการเหมือนต้นเดิม ดังนั้นอาการโรคของวิสา strain ใหม่นี้จะเห็นได้ก็ต่อเมื่อวิสานั้นถูกแยกและถ่ายทอดไปสู่พืชอาศัยเฉพาะต่างหาก การเปลี่ยนแปลงของวิสาได้ strain ใหม่นี้เกิดจาก mutation เหมือนจุลินทรีย์อื่นๆ
ความแตกต่างของ strain ของวิสาอยู่ที่คุณสมบัติเฉพาะซึ่งเกี่ยวโดยตรงต่อความสามารถของวิสา ในการทำให้เกิดโรค สังเกตได้จากวิธีการติดเชื้อ อาการโรค ความเสียหายของโรค ผลิตผล อุณหภูมิที่เหมาะสมต่อการเจริญของโรค อัตราการเคลื่อนย้ายของวิสาในเนื้อเยื่อพืชอาศัย พืชอาศัยต่างๆ แมลงพาหะ รวมทั้งคุณสมบัติของน้ำคั้นจากพืชที่เป็นโรค เช่นอายุของเชื้อในหลอดเก็บอุณหภูมิห้องที่ยังก่อโรคได้ อุณหภูมิความร้อนที่เชื้อหมดสภาพก่อโรค และจุดที่วิสาในนํ้าคั้นเจือจางที่สุดที่ยังสามารถทวีจำนวนได้ในพืชอาศัย เป็นต้น
ไส้เดือนฝอย
ไส้เดือนฝอยศัตรูพืชหลาย species และ genera ที่มีความแตกต่างกันทาง races เช่นไส้เดือนฝอยพวก cyst (Heterodera spp.)ไส้เดือนฝอยรากปม (Meloidogyne spp.) ไส้เดือนฝอยที่ลำต้นและหัว(Ditylenchus dipsaci) และไส้เดือนฝอย burrowing (Radopholus similis) Races ประกอบด้วย biotypes จำนวนหนึ่งของชนิดที่มีลักษณะต่างๆ ในความสามารถที่ทำให้เกิดโรคเหมือนกัน การเปลี่ยนแปลงที่เกิด race นี้เกิดได้จากการ hybridization และ mutation ซึ่งจะมีผลทำให้มีการเปลี่ยนแปลงพืชอาศัยต่างๆ ขึ้น
ที่มา:ไพโรจน์ จ๋วงพานิช
